
Ecosystem‐Level Conservation
We now turn to conservation practices, beginning with ecosystem-level approaches. The fundamental goal of conservation does not change under climate change—it is still to protect biotic systems from the deleterious effects of human activities. Therefore, the conservation tools we discussed in Chapters 6–8 remain relevant and are applied largely for the same purposes. The main change, in terms of working objectives, is that we must now also ensure that climate-induced ecological transitions unfold as they would in undisturbed systems. This entails facilitating species adaptation.
There are four main concerns related to adaptation that warrant management intervention:
1. The natural capacity for adaptation of many species has been compromised by human activities, making them less able to withstand climatic variability and less able to shift their range
2. Barriers to movement now exist, including regions where habitat quality has deteriorated
3. This episode of warming is particularly rapid and may exceed the intrinsic adaptive capacity of some species, especially if the warmest scenarios are realized
4. Non-native species may competitively exclude the movement of native species into new areas
In terms of implementation, adapting conservation to a changing climate is mainly a matter of fitting existing conservation methods into a dynamic framework (Lindenmayer et al. 2010; Abrahms et al. 2017). No truly novel methods exist for addressing climate change, though some existing approaches may be used in new ways.
Protected Areas
The need to provide species with a refuge from anthropogenic disturbances does not diminish under climate change. In fact, the need is greater. Protected areas increase the resilience and adaptive capacity of species, thereby supporting climate adaptation (Hodgson et al. 2009). In addition, protected areas enhance landscape connectivity and serve as ecological benchmarks—roles that take on added importance under climate change.
Because the distribution of species and ecosystems is destined to shift, reserve planning now requires a dynamic framework (Game et al. 2011). Rather than trying to protect specific assemblages in specific locations, we should aim to provide species with continual access to protection as their range shifts across the landscape. This is best accomplished by representing the stable geophysical attributes that underlie ecological patterns (Anderson and Ferree 2010; Groves et al. 2012). Simply put, our objective should be to represent the major “arenas” of biological diversity, not the “actors” (Beier and Brost 2010). Bioclimatic modelling suggests that this approach should succeed in maintaining coarse-filter representation as long as movement among reserves is not overly restricted (Schneider and Bayne 2015).
A key challenge in implementing the arenas concept is identifying the arenas (Schneider and Bayne 2015). What is the appropriate scale? Which geophysical attributes should be used? Are some attributes more important than others? And how do we delineate features that blend seamlessly from one to the next? These are difficult questions to answer from a purely abiotic perspective. Therefore, we must look to ecological patterns for guidance. The geophysical features that matter are those that have a demonstrable ecological effect.
A hybrid system, such as the National Ecological Framework, is well suited to this task (Fig. 9.12). This classification is fundamentally based on broad geophysical features and patterns. However, vegetation patterns are incorporated as well, providing insight into the ecological relevance of the observed geophysical features and indicating where distinctions are warranted. There is no guarantee that the ecosystem boundaries in this classification will all remain stable under climate change. But this approach should be robust enough to ensure that the major biotic arenas have been represented.
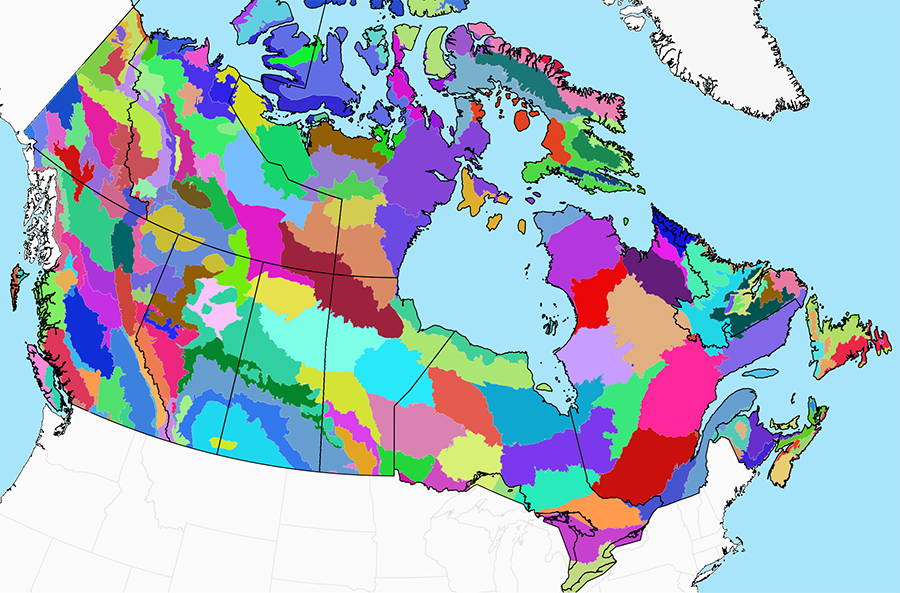
Finer-scale landscape features can be represented through “land facets,” which are landscape units with uniform topographic, geologic, and soil attributes (Wessels et al. 1999; Beier and Brost 2010). For example, land facets might include marshes, patches of sandy soil, riparian zones, and other features relevant to the region. In contrast to the large polygons of the National Ecological Framework, which are all unique and merit individual representation, land facets are recurring landscape features. The intent is not to represent each facet individually, but to capture a sample of each type within each ecoregion.
At present, there is no established classification system for land facets in Canada. Therefore, the task of defining them falls to protected area planning teams. As previously noted, it can be challenging to determine which features to include and how to set breakpoints. As a general rule, features should be selected because of their ecological importance, not just because the GIS layers happen to be available.
Climate Refugia
A new dimension to reserve design that has emerged in response to climate change is the protection of climate refugia, of which there are various types (Ashcroft et al. 2012; Morelli et al. 2016). One type, termed in situ refugia, involves landscapes that are expected to be relatively resistant to ecological change (Rose and Burton 2009). For example, a 2°C rise in temperature in a mountainous area represents only a few hundred metres of vertical rise (Loarie et al. 2009). A single vertically oriented reserve in such an area could serve as a climate refugium because it could maintain representation of most pre-existing ecosystem types, even if they shift upslope under moderate levels of warming. In contrast, a 2°C rise in temperature in a region with little topographic relief may generate a northward displacement of several hundred kilometres, precluding comprehensive protection.
The northern fringe of large ecosystems may also serve as in situ refugia, since these areas will be among the last to transition. However, such refugia should not be considered permanent, even if bioclimatic projections suggest they will remain stable. Climate change will continue until well into the next century, beyond the scope of our models. Few, if any, ecosystem remnants are expected to remain in place indefinitely. What we are identifying are areas of relative stability, which is still important for conservation. Protecting these areas may provide species with the time they need to adjust to the changing climate.
As with other reserve design elements, the effects of including in situ refugia in a reserve network should be explored with conservation planning software. Trade-offs with other design objectives can be expected. Because refugia are based on climate model projections, the level of uncertainty is high, and this will weigh against them in trade-off analyses
Ex situ climate refugia are also possible. The idea here is to protect the projected future location of selected ecosystem types. This might be proposed for ecosystems that have high ecological or social value, or that are expected to become rare in the future. This approach is technically feasible, but the selection of such sites may be difficult to defend. Why should some ecosystem types be afforded ex situ protection over others? Which point in time should be targeted? How much uncertainty about future projections is acceptable? These questions are difficult to answer objectively.
Climate refugia can also be identified at scales finer than those normally used for protected area planning. These micro-refugia are local sites of ecological stability that persist, at least for a time, after the regional climate envelope has transitioned to something new (Dobrowski 2011; Lenoir et al. 2017). Their existence is a consequence of topographic complexity and associated fine-scale climatic patterns. Because micro-refugia are small and diffuse, they do not lend themselves to protection through conventional reserves. However, their temporary protection through special management zones may provide many species the extra time they need for climate adaptation.
Connectivity
Climate change greatly increases the need for landscape connectivity, to support the ability of animals and plants to track their preferred climatic conditions at the necessary pace. The management tools for enhancing connectivity described in earlier chapters (i.e., generic connectivity measures, movement corridors, and protected areas) remain the mainstay approaches under climate change. No new options are available.
Generic connectivity measures facilitate movement in all directions and are therefore robust under climate change. Because they apply to the entire land base, these measures are well suited to facilitating range shifts and enhancing the viability and adaptive capacity of species (Doerr et al. 2011). The only change needed to accommodate climate change is a redoubling of efforts.
Corridors facilitate movement in specific directions, so their effectiveness is affected by the directionality of movement imposed by climate change (Littlefield et al. 2017). This needs to be accounted for at the design stage (Beier 2012; Nunez et al. 2013). For most species, range shifts will occur along existing climatic gradients, typically northward or upslope. Planners should assess corridor options on the basis of their alignment with such gradients, favouring those that are most aligned (Rouget et al. 2006; Nunez et al. 2013).
Protected areas present a special case in that they contribute to landscape connectivity while also depending on connectivity to remain functionally linked. Both aspects need to be considered in the reserve design process, with special attention given to the effects of climate change.
Connectivity among reserves can be maximized at the planning stage by taking advantage of the directionality that climate change imposes on species range shifts. For example, instead of creating a circular reserve, it would be advantageous to create an oblong reserve oriented in the direction that climate envelopes are expected to move. Connectivity can also be enhanced across the entire reserve network by arranging large reserves and smaller “stepping stone” reserves in a configuration that minimizes the distance between reserves along a relevant climatic gradient (Fig. 9.13; Robillard et al. 2015).
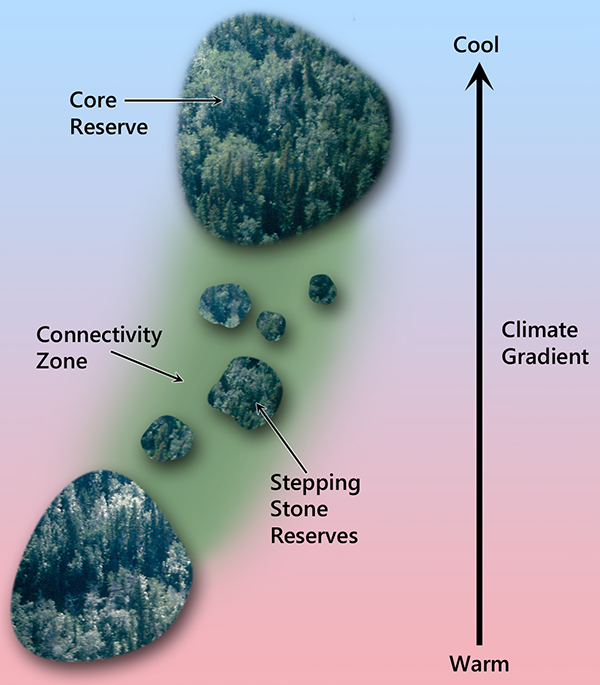 |
Fig. 9.13. Long-distance range shifts can be facilitated by aligning large core reserves, stepping stone reserves, and special management zones with the directionality imposed by climate change. |
Design approaches for enhancing connectivity among reserves are subject to several practical limitations. In particular, we are not working with a blank slate; more than 12% of Canada has already been protected. This places constraints on the overall reserve system configuration, limiting options for minimizing distances along the axes preferred for accommodating climate change. Also, the incorporation of preferred spatial axes is not something that conservation planning software was designed to do. Time-consuming work-arounds and manual approaches are required. Finally, the distances between existing reserves are often vast and not easily bridged.
The placement of new reserves along preferred climatic axes may also conflict with objectives related to representation, since the respective spatial priorities are unlikely to align just by chance. When conflict is unavoidable, planners should try to find a workable balance, rather than emphasize one class of design objectives over another. For example, it may be necessary to accept gaps in fine-scale representation to obtain the flexibility needed to achieve functional objectives like connectivity.
The Natural Disturbance Model
The natural disturbance model is a method for reducing proximate threats to biodiversity from industrial disturbances on the working land base. It remains an important conservation tool under climate change, helping to prevent species declines and enhancing resilience and adaptive capacity.
The natural disturbance model is not intrinsically forward-looking, so its application does not depend on climate projections or bioclimatic modelling. The intent is simply to modify industrial practices such that ecological composition, structure, and processes remain within the natural range of variation (NRV). Estimates of NRV that are based on historical conditions remain acceptable for now because climate-induced ecological transitions are not yet widespread (Gauthier et al. 2023). In the future, a dynamic ecological baseline will be needed.
A dynamic version of NRV can be obtained by monitoring relatively undisturbed landscapes and then extrapolating the findings to the working landscape. As undisturbed sites evolve under warming temperatures, so too will NRV. The larger sites in our existing system of protected areas are obvious choices as benchmarks; indeed, this is already one of their central roles (Wiersma 2005). However, significant gaps in representation exist in many regions. To some extent, these gaps can be addressed by monitoring smaller natural areas and sites that are only partially protected. But it would be best to fill gaps in representation by adding new protected areas, adding impetus to Canada’s commitment to protect 30% of its landscapes. To maximize their effectiveness as benchmarks, the new sites should be large enough to maintain and represent natural ecological processes.
Box 9.3. A Functional Approach for Defining Ecological Baselines
To accommodate climate change, some researchers have proposed that ecological baselines should be based on ecosystem function, rather than specific structural and compositional attributes (Hagerman et al. 2010b; Glick et al. 2011). For example, it has been suggested that we maintain “functional” ecosystems and ecosystem “health.” The appeal of these sorts of measures is that they obviate the need for periodically updating the ecological reference state.
The problem with this approach is that objectives like maintaining ecosystem function and health are inherently ambiguous and malleable. We achieve robustness at the expense of clarity. Furthermore, the relationship between ecosystem function and species diversity is quite tenuous (Srivastava and Vellend 2005). The loss of individual species, especially those that are rare, is unlikely to be detected by any of the available measures of ecosystem function (Schwartz et al. 2000).
Given the malleability of functional objectives and the lack of an explicit connection to species diversity, ecological baselines defined solely on the basis of functional attributes provide a poor foundation for biodiversity conservation. The functional approach is best restricted to ecosystem services applications. For conservation applications, an integrated approach that includes structure, composition, and specified functions, explicitly linked to natural systems, is best.
Restoration and Reclamation
Like the other ecosystem-level methods we have discussed, restoration and reclamation also need to be placed in a dynamic framework. Since most of these projects are focused on the restoration of vegetation, the main issue is determining the appropriate source of seed stock.
One line of reasoning suggests that seed stock should be obtained from warmer sites along a relevant climatic gradient. The idea is to preadapt restored sites to future climates to maximize their long-term stability (Harris et al. 2006). This approach has gained significant support in the forestry sector. Regulations requiring the use of local seed stock in reforestation programs are now being modified to permit the use of seed from warmer climatic zones (Klenk 2015).
An alternative view is that we should continue using local sites as seed sources. In this case, the conservation of local genetic adaptations is the overriding concern, not the climate-proofing of restored sites (Gibson et al. 2009). Although local populations may not be well suited to future climates at their current location, their genetic adaptations will be important in facilitating range shifts into cooler landscapes in future decades (Gibson et al. 2009; Hamann and Aitken 2013).
Both of these perspectives have merit and the choice of which seed source to use depends on the project objectives and local context. For example, restoration projects in agricultural landscapes may emphasize local seed sources, given the importance of conserving local genotypes when little native vegetation remains. Conversely, reclamation projects in northern forests may emphasize preadaptation, given the slow growth and longevity of trees. A proviso when using stock from warmer sites is that the stock must be sufficiently winter hardy to survive in the restoration site under current climate conditions. There are also regulatory issues that must be addressed if the seed stock is coming from the US.
Invasive Species
Climate change will make Canada’s harsh climate more hospitable to alien species, adding a new dimension to invasive species control (Smith et al. 2012). Our ability to achieve successful outcomes will decline, and costs will rise, suggesting that the cost-benefit analyses underlying current control programs will need to be revisited. In short, despite increasing need, we have to be even more selective about the programs we undertake.
We will also need to reconsider how alien species are identified. Until now, the introduction of alien species was largely mediated by humans, and their status as non-native was rarely in doubt. In the future, many new arrivals will be species from the northern US moving into Canada on their own in response to climate change (McKenney et al. 2007). This confers considerable ambiguity as to their status.

Under the conceptual framework developed earlier in the chapter, the coming “invasion” of northern US species should be accepted as a natural ecological response to warming temperatures (Walther et al. 2009). These northward-moving species will have an essential role in populating the new climate envelopes that will develop in Canada’s southern landscapes, filling niches that our native species are poorly adapted to (Berteaux et al. 2010). Consider the migration of buffalograss. This is a warm-season grass that is widespread in the US Great Plains but rare in Canada (Fig. 9.14). Under warmer and drier conditions, this species will fill new dry grassland niches in Canada that existing species are not well suited to, thereby helping to maintain ecosystem stability and function.
Major ecological disruptions are unlikely to arise from northern US migrants (Mueller and Hellmann 2008). These species will be at the northern edge of their range and poorly adapted to cooler Canadian environments (otherwise, they would already be here). Moreover, they will be moving in response to climatic changes rather than invasive tendencies. Nevertheless, exceptions may occur, forcing managers to contemplate control measures. In these cases, it will be the potential for economic and ecological disruption, rather than simply their alien status, that is most relevant (Coops et al. 2008).
Box 9.4. Alien Species as Endangered Species
Under the Species at Risk Act, a species must have extended its range into Canada without human intervention and have been present for at least 50 years before it can be recognized under the Act (GOC 2002, Sec. 2). The accommodation of climate change demands a more nuanced approach (Gibson et al. 2009). Species that are at risk in the US should not be ignored for 50 years after they establish in Canada just because they are not historically “ours.” On the other hand, it would be inappropriate to list all incoming species as threatened simply because these vanguard populations are inherently unstable. A dynamic and integrated approach is needed.
